千島学説|新生命医学会
千島学説|新生命医学会
(a) 脾臓の構造
脾臓は多量の血液を含む暗赤色の扁楕円形の臓器で、胃底の左後方に位置しています。表面は結合組織と平滑筋から成る被膜で被われています。脾臓は平滑筋がリズミカルな伸縮運動をすることから『腹の心臓』とも呼ばれています。脾臓内の動脈周囲には鞘状になった白色脾髄といわれるリンパ組織があり、他に静脈周囲の赤色脾髄があります。特に興味深いことは白色脾髄は動脈と、赤色脾髄は静脈と密接に結びついていることです。血管や神経は脾門を通り、動脈は白色脾髄の中央部を通って中心動脈になっています。白色脾髄に多くの毛細血管を与え、その本管は次に赤色脾髄へ出てから筆の穂先状に分かれた毛細血管になります。さて、そこで問題となるのが脾の働きは何か、そして細小動脈の先端は閉鎖型か開放型かということです。
(b) 脾臓の働きと赤血球との関連
ギャレアスは脾は働きが解らない不思議な器官だといいましたが、今日になってもその働きが殆ど解明されることなく経過しているようです。脾の働きについての研究は主として摘出手術の方策や、脾の移植実験、血管の結窄よる事後経過などが行われてきました。それらの実験結果から大体次のような働きが脾臓にあるといわれています。
① 赤血球の造血作用……これについて次のような報告があります。『胎生5ケ月までは脾は間葉細胞→赤血球母細胞→赤血球の過程をとるが、6ケ月以降はこの働きが止まってしまう』 この過程は実際の状態を逆に見ているようです。赤血球が脾細胞に分化しているのが通常で、脾の間葉細胞が赤血球を造るのではありません。クロスビーもこの脾臓造血説を支持していますが、成体での脾造血は飢餓或いは栄養不良時の場合のみ脾細胞が赤血球に逆分化するだけです。これまでの研究者のすべてが脾の造血作用を報告するとき、実験動物の栄養状態を正常状態に保つ注意を払っていません。このため飢餓或いは栄養不良になった病的状態の実験動物を使用するために、正常時とは逆の脾細胞から赤血球へ分化する過程を見て脾造血を唱えているのです。動物の栄養不良時には赤血球の分化過程が逆転するという可逆的分化の実際を知らないために起きる誤りといえます。
② 赤血球造血の調節……フェレータは脾臓は正常体では骨髄の赤血球造血を抑制する働きがあると考えていて、これは脾の働きが過度になると骨髄の造血機能が退行することが認められるからだと説明しています。これに対し千島喜久男は次のように批判しています。
『慢性貧血の場合には、骨髄脂肪から赤血球への逆分化は殆ど停止し、次に他組織からの逆分化に移るがこの骨髄脂肪からの分化停止状態を退行と見たのだろう。骨髄の状態に対し、脾においては病的性質をもった赤血球の脾細胞への分化によって脾が著しく腫大する事実によって見誤った解釈をしている。脾が栄養状態に応じて行う間接的な赤血球造血の調節作用は確かだが、骨髄と脾とが造血に関して相反する作用を示すのは病的といえる脾の貧血性腫大が起きたときに限る。正常体においては程度の差はあるが、すべての組織と赤血球との栄養の変化による可逆性は同一傾向を示すものだ』
③ 赤血球の表面積と脾摘出……赤血球の表面は静電気的な保護膜で被われていますが、赤血球が老化すると静電気過重荷電によって粘着性が低下するといわれています。脾臓を摘出した動物の赤血球は扁平化してその表面積は大きくなるが容積には変化がないという研究もあります。しかしこれは脾の重要な働きと関連があるようには思えません。
④ 赤血球の貯留作用……脾臓は著しい拡張性をもっていて、その赤色髄や静脈洞内には多量の赤血球を貯え、闘争時や走る前に必要に応じてそれを血管内へ戻す機能があるとバークロフトはいっています。しかしこのことは、人間の場合には余り重要なこととは思えません。なぜなら人間の脾臓には平均して30~40ミリリットルの赤血球は貯えていますが、病的な腫大を除き拡張性はもっていません。脾臓の被膜に筋肉線維をもっていないからです。脾に赤血球を貯える働きがあることは否定できませんが、それが生命に重要な関与をしているものでないことは、脾臓の摘出をしても生命現象には大きな影響がないことからも解ります。
マッケンジーも脾臓は赤血球の貯蔵場所ではないといっています。それは絶食させたマウスの脾が重量を3分の1に減らし、赤血球も減少していたことを理由としています。結論はともかくとして、絶食させた、いわゆる病的動物の実験を正常体にも適用したことは正しいとはいえません。
⑤ 血球の選別作用……クロスビーは『脾臓はそこを通過する赤血球を選別する働きがあり、骨髄で造られたもののなかから形や性質が正常でない約10%の赤血球を抑留する。それらの赤血球は血管内の喰細胞によって捕喰される』と報告しています。彼は遺伝性と考える球形赤血球症患者の血液を正常体の人に輸血すると、15日以内に球形赤血球が消失するのに対し、脾臓を摘出された人に輸血すると球形赤血球は殆どが、正常な寿命とされる120日を保つことを主張の理由に挙げています。このような作用があることを否定することはできませんが、血管内の喰細胞によって抑留赤血球が捕喰されるという説には、そのような細胞は存在しないという事実から賛成できません。
⑥ 鉄代謝と赤血球破壊の場所……ポンフイックが1869年、始めてこの説を唱えてから、〈脾は赤血球を破壊する場所である〉と多くの賛同者を得て一般的な常識のようになってしまいました。また赤血球のヘモグロビンが解体することによりその鉄が遊離し、赤芽球の形成に貢献するというクロスビーの意見もあります。しかし赤血球が固定組織細胞に分化するときにはヘモグロビン中の鉄分はその組織で吸収され再び利用されることはすべての組織についていえることで、鉄代謝は脾に限ったことではなく、脾の特質といえる作用ではないといえます。
⑦ 赤血球の細胞質の異物除去……クロスビーは『脾は赤血球の細胞質中にある粒子の如き固形体を除去する働きをもっている』と唱えています。それは溶血性貧血患者に見られる含顆粒赤血球を含む血液を正常体に輸血すると約4時間後には顆粒の殆どが消滅するが、脾臓摘出を施した個体ではなお80%前後が残留していたことから、脾は赤血球の細胞質から鉄顆粒を除去する働きをもつと主張したわけです。このクロスビーの説は一見すると尤もらしい説ですが、生物学的に正しい認識をもっている研究者には賛同されないと思います。なぜなら、細胞体を破壊することなく細胞質内の固形体である異物、いわゆる鉄顆粒だけを除去するといったマジック的な体機能は、化学的、形態学的に過程を確認されていませんし、またそれは不可能なことだと思います。鉄を含む細胞の浄化は脾に限定されるのではなく、体の組織のすべての箇所で行われていることは、赤血球の分化能を説明する千島学説の第1原理・赤血球分化説から理解できることでしょう。
⑧ 老化赤血球の処理……ルースやロバートソンが1917年に唱えた『老化した赤血球は脾で細かく破壊されてその一生を終える』という悲劇的な説が現在の定説になっています。確かにこのような状態が見られることは正常体においても千島喜久男は認めています。しかし見方が従来の研究者とは違っているのです。千島はこの状態について次のように説明しています。
『従来の説のように、赤血球がこのように脾で破壊され消滅していると考えることに大きな誤謬がある。哺乳類の無核赤血球は、如何にも壊れ去っているかのように考えられているが決してそうではなく、脾に限らず殆どの体組織で赤血球が固定組織細胞に分化する際には一度、融合により赤血球の形を崩し、いわゆる〈赤血球モネラ〉を形成し、その内部に核を新生するが、多数の赤血球塊から生じたモネラ中には幾つかのリンパ球に似た核を新生する。この像は赤血球が破壊されているのではなく、赤血球が細胞に分化する移行途上の状態である。赤血球が塊の状態で又は単独で核のある固定組織細胞に分化する途上に見られる赤血球の融合、いわゆるモネラ状態は脾に限ったことではなく体組織全体に見られるものであることを忘れてはならない。栄養不良のときはこの逆の過程が生じ、組織細胞から同じような過程を経て赤血球に戻る。この逆分化の詳細は別の機会に譲る。脾が異物や病的赤血球を抑留する作用があることは否定できない。肝と共通した作用といえる』
マキシモウは『人間の脾の赤色髄中には貪喰細胞があり、主として赤血球、時には異物を貪喰しており、その消化段階によって黄色や褐色の顆粒を含み、鉄反応を示すものもある』といっています。 それよりずっと以前にキースがニワトリの脾の赤色髄中に赤血球貪喰細胞があると報告していました。ハンスもウサギの脾に赤血球貪喰細胞を見たといっています。しかし、これらの報告は確かに像は真実そのように見えても、貪喰細胞なるものが予め血管内に存在して赤血球を捕喰したのではなく赤血球の融合状態を貪喰細胞が赤血球を捕喰しているように見えて見誤ったのでしょう。
(c) 脾臓と血管との特殊な関連
脾臓の血管系は特殊なものであることは古来から知られていました。緒方知三郎氏は『今、仮に私が脾臓を始めて発見し、その名付け親になったとしたら、私はその構造にふさわしい血液腺とか血液器という名を与えただろう。脾の血液に対する関係がリンパ腺のリンパ液に対するそれと同じだからである』といっていますが、さすがといえる着想です。
脾は前述したように古来から赤血球やリンパ球の生産、老化赤血球の破壊、異物や毒物の抑留、血液の貯蔵場所等々といった働きをこなす場所と考えられているのも、脾と血液の密接な関連があることを示しているといえます。
(d) 脾の毛細血管は開放型か閉鎖型か
これまで毛細血管は血管内皮細胞に被われ、その末端部は完全に閉鎖型であるとされていますが、脾臓の血管系だけそれとは異なり3つの説に分かれて今日でも論争が続いています。
① 開放型説……静脈に注入した物質や輸血した血液の赤血球が脾の実質組織中の毛細血管外に数多く散乱している事実は殆どの組織学者が認めており、脾の毛細血管は開放型だと主張する説です。
トーマス、ロビンソンたちは細小動脈が脾組織中に開放していることを確認し、モールは拡張した脾において毛細血管は開放型だと唱え、マッケンジーはネズミの脾について観察した結果、開放型であると結論しました。開放型説は事実を見ている研究者の説であり最も妥当な説だといえます。
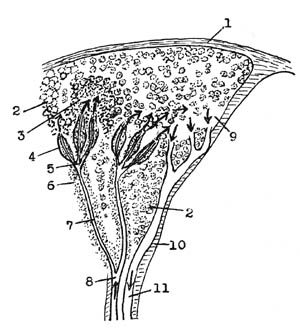
〈脾の血管の末端は開放型である〉
原図・千島喜久男
1…皮膜 2…赤色脾髄
3…白色脾髄 4…莢動脈
5…筆毛動脈 6…リンパ球集団
7…中心動脈 8…脾動脈の分岐
9…脾洞 10…脾梁
11…脾梁静脈
※矢印は血液の流れを示します。
② 閉鎖型説……スヌークはモルモットの脾の網状組織を、ヘリンガーは人間の脾で、ドーゲットはイヌの脾に薬剤を静脈注射して観察した結果、何れも細小動脈と静脈洞とは連続した内皮細胞に被われながら連結しており閉鎖型になっていると主張しています。
エリアスは脾の毛細血管はその壁が明瞭であるにも拘わらず赤血球が管外組織に出ていることを認めていますが、これは観察方法や実験材料のミスによるものと考え閉鎖型説に近い考えをもっています。傾向として閉鎖型説を唱える研究者は事実を曲げて既成説に同調する姿勢が窺えます。
③ 折衷説……開放型を示す部分と閉鎖型の部分との両方があるという説で、ドーゲットは最近では大部分の研究者がこの折衷説を支持しているといっています。
④ 脾静脈洞の濾過説……脾は滑平筋によって自動的にリズミカルな伸縮運動をしていることは生体でも実証されています。この運動は血圧や呼吸とは無関係ですが脾への神経の一部を切断すると止まります。ネルやモールたちは脾動脈中へ酵母やアスファルト粒子を注入し観察した結果、大きい粒子は静脈洞内に抑留されているのを見て、静脈洞壁がフィルターの役目をして血漿だけは通過させるが赤血球は残留させる作用があると結論しました。これはドーゲットがその著のなかで伝えたものです。 このネルたちの報告にドーゲットも賛同していますが、これは想像説だと千島喜久男は批判し次のように説明しています。
『血管端末の開放型説の主張者は、脾の髄質中には常に多数の赤血球が固定細胞の隙間に不規則に散乱しているのを見ても、この部分で赤血球が生産されているような証拠はまったく認められないから、これらの赤血球は流血中のものが血管壁の開放部から流出したものだと判断している。この考えは正しいものだ。一方、閉鎖型説を支持する人たちは、この脾髄質中にある赤血球は通常赤血球より小さいから、血管端末から流出したものとは考えられず、もし開放型だったら脾の赤色脾髄は赤血球で充満されなければならない。しかも白色脾髄もあるのだから開放型説は間違っていると主張しているが、この主張は全く間違っている。なぜなら、第一に赤色脾髄中の赤血球はその多くが通常赤血球と同じ大きさであり、染色性ももっている。見たところでは確かに正常赤血球より小さいものもあるが、これは赤血球が血流の停止によって球形化したあと、リンパ球に変わる前段階として脾に限らず全ての組織で広く認められる現象である。第二に、開放型なら赤色脾髄は赤血球で充満するはずだという主張は赤血球を固定的なものと観ていることからくる誤った判断である。
赤血球→球形化→リンパ球へと分化し、そして脾細胞のみならず全ての体細胞へ分化する潜在能力をもっているから、脾の実質中には赤血球が流入してから経過した時間の差によって、また赤血球が存在する組織の電気的誘導作用によって分化方向も異なることになるから、赤色脾髄が赤血球で充たされねばならぬという理由は少しもない。開放型であっても種々の細胞があったり、白色脾髄があることは赤血球の分化能から当然のことである。閉鎖型説を唱える人々に共通することは固定的、且つ機械論的な考えをもっていることである』
⑤ 脾の生体観察……脾の血管系問題を解明しようとして生きた状態の脾を観察する試みがなされています。マキシモウやブルームの報告では、ある研究者たちは生体の脾の血流は著しく間歇的であり、静脈洞内の血液の液状成分は血管壁で濾過されたあと壁外に出ることはある。しかし血管は原則として閉鎖型だと主張しています。また別の研究者たちは生きた脾の観察から、脾の血管は開放型になっており血液は細小動脈の端末から直接に脾組織細胞の隙間に流出する状態が確認できるといっています。そして細胞の間隙に出た赤血球はそこに蓄えられ、また赤血球と血漿が分離するのも、静脈洞内ではなく血管から流出した組織細胞の間隙だとしています。マキシモウとブルームは現在までの諸観察を総合して考察すると開放型説に有利な証拠が多いと開放型説に軍配を挙げた形をとっているようですが、これは妥当な判断だといえます。これに対してドーゲットは『アメリカでは閉鎖型だとするのが常識だ』と、あくまでも閉鎖型説にこだわっていますが、その前に、いま一度、組織間隙へ流出する血液の状態を注意深く観察してほしいものです。
老廃物を体外へ排泄する器官は動物の種類によって種々変わりますが、代謝産物には共通したところがあります。アメーバは老廃物を尿素として排泄し、高等動物でも尿素や尿酸、またアンモニアとして排泄しています。この他に水分調節作用もまた排泄器官の大きな役割です。
生殖器官は発生学的に排泄器官(中腎)の一部が分化して生じた、一種の排泄器官と観るべきだと千島喜久男はいっています。
アメーバやゾウリムシ、その他多くの原生動物の細胞中にある伸縮砲といわれるものは、リズミカルな伸縮によって老廃物を体外へ排出するといわれていますが、これは排泄器官である一方、循環器機能やさらに呼吸などの作用を助ける機能も併せてもっていると考えるのはどうでしょうか。
呼吸、循環、排泄は高等動物では密接な関連性をもっているのに、原生動物においてはこの3系統がはっきり分化していません。その点から原生動物の伸縮砲がこれらに関わっているのではないかと推測されるわけです。
(a) 脊椎動物の腎臓
脊椎動物の腎臓には大体次の3型があります。
①前腎……胚子の腎で最も原始的なものです。下等な魚類ではこの前腎が生体でも機能します。
②中腎……前腎より複雑な管の集まりから成っています。高等な魚類や両棲類では一般に、発生の途中で前腎が退行し、中腎が生体の排泄器官になります。
③後腎……後腎になると複雑化し、前腎、中腎などより後に発生します。ボウマン氏嚢、糸毬体、尿細管などの分化も明瞭になります。爬虫類、鳥類、哺乳類の生体の排泄器官として作用します。
千島のニワトリの胚子による観察では、中腎の前縁部分から副腎が発生し、内縁からは生殖巣が発生します。中腎の一部は副腎や生殖巣の組織に分化し、また一部は細胞の崩壊などによって退行し、孵化時には中腎は痕跡を残すだけになります。一般に人間や他の哺乳類の腎臓と呼んでいるのは後腎を意味します。
腎臓は糸毬体や尿細管の間隙などに血液が充満していること、また発生の初期には後部大動脈の一分枝から血液が流出して生じた赤血球塊から、リンパ球状過程を経て腎臓の原基ができることを、世界で初めて発見したのが千島喜久男でした。
(b) 腎臓の構造と機能
爬虫類以上の成体では前にもいいましたように、一般に後腎を腎臓といいます。腎の皮質は腎小体とそれから出る細長い尿細管があります。腎小体は毛細管から成る糸毬体とこれを包むボウマン氏嚢とから組み合わされています。腎小体は従来の書籍には模式的に進入細動脈と排出細動脈及び毛細管網がはっきり示されていますが、実際には糸毬体の毛細管は意外にはっきりしていません。
尿は糸毬体と尿細管で造られますが、その機構については濾過説、分泌説などがあってまだ解明されていない現況です。腎は腎動脈から入る血液によって灌漑され、また交感神経、副交感神経の強い支配下にあり、自律神経の不調和に対し敏感に反応し機能にも乱れを生じさせます。肺は血液浄化のために空気を媒体としますが、腎は血液成分の浄化を行うという働きから考えると血液の浄化という意味において肺との共通点をもっています。
(c) 腎の組織発生
1個の腎小体とそれから出る尿細管を合わせてネフロン(腎単位)と呼ばれていますが、これと尿細管が集まって作られた集合管とは別々の起源によるものといわれています。
つまり腎単位は間葉性細胞様の細胞塊からの分化によって生じ、集合管は中腎管から外部成長によって形成されるというマキシモウらの主張が通説となっているわけです。しかし千島喜久男がニワトリの胚子で研究した結果、実際は腎の原基は後大動脈の分岐から腎形成の場所へ血液が流入し、その赤血球塊が腎原基の間葉性細胞に分化し、それを基礎として糸毬体や尿細管が形成され、そのあと既成尿細管の間隙へ次々と流入する赤血球の集塊から新しい尿細管が形成されていくことを確認しています。
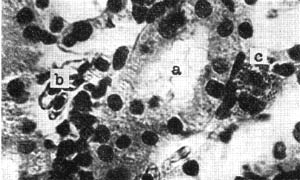
写真はオタマジャクシ(体長60ミリ)の腎。
尿細管(a)の間隙に赤血球(b)及びエオジン好性顆粒白血球(c)があり、これらが尿細管壁の細胞へ移行する過程を示している。
(撮影・千島、鈴木)
この事実は精巣における精細管の増加が生じる過程と同じです。終生において細管が存在するのではなく、一定期間の経過後には退化し次の新しい細管の形成によって補充されていきます。
