千島学説|新生命医学会
千島学説|新生命医学会
① 生殖細胞と原生生物との類似
細胞の起源と微生物との共生現象をお話するまえに、まず生物体中で最も根本的な存在である赤血球と生殖細胞とが原生生物その他の下等生物に似た性格をもっていることを述べておきたいと思います。生殖細胞は非常に原始的な形質をもっていることは周知の事実となっています。
卵子はアメーバの休止期に、精子は鞭毛虫或いは帽針状腐敗菌に似た形をもっており、生殖細胞は分化した多細胞生物が再び原始の状態に戻ったものと解することができます。
このように考えるとき、生殖細胞と下等微生物との形態や習性に類似点があることは単なる偶然の一致ではなくて、生殖細胞は多細胞生物発生という原始状態に戻り、過去の歴史を反復する段階にあるものということもできる筈です。
② 赤血球と原生生物との類似
現代の血液学では赤血球は最高度に分化した細胞だと定義づけしています。このことについて千島喜久男は『赤血球は生殖細胞より一層に原始的で細胞以前のもの』といっています。
赤血球がまだ原生生物的形質を多分にもっているという証拠は、カバースライド法で両棲類、鳥類や哺乳類などの生きた赤血球を観察することで理解できることでしょう。
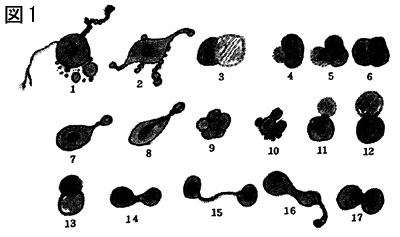

(1) カエルの赤血球は一部が細長く延長し、その先端は鞭毛状になって緩やかな鞭毛運動を示します(図1)。また赤血球の表面にはしばしば鞭毛状の突起を生ずることがあります。ニワトリやウサギ、ヒトといった哺乳類の赤血球を体外に取り出すと、ときに飴の金平糖のように変化します(図2)。浸透圧が異なった状態が加わるとこのような変化は一層はっきりしますが、全血液そのままでも往々にしてこのような現象を見ることができます。突起は鞭毛虫の鞭毛に相当するものだと千島は述べています。ミンティンは原生生物の繊毛は進化論的に鞭毛に先行するものだといっていますが、金平糖状赤血球の突起が鞭毛に該当し、アメーバ状運動をする白血球はその後から現れるという千島の観察はミンティンの説にも合致しているようです。しかも、このような変化は血管内を流れている赤血球では生じません。血流の停止、或いは体外に取り出したとき始めて見られる現象です。しかし、オタマジャクシの尾部毛細管を生きたまま観察していると、赤血球が毛細管壁を通過する際に、鞭毛状の突起を出して管壁を穿孔し赤血球内容がこの小さな孔を通って血管外に出ると、それは白血球に変わるという不思議な現象を見せてくれます。
赤血球が原生生物に似た行動を示すのは、赤血球が白血球に分化する途中及び白血球に変わってからです。
(2) 各種動物の赤血球は始め円盤状ですが血流が停止すると同時に球形に姿を変えます。これは卵子やアメーバの保護嚢に似て一種の原始状態への復帰と考えられます。哺乳類の赤血球の球形化と金平糖状変化はあい伴って起きます。
(3) 白血球が偽足を出してバクテリアその他を貪喰するかのような行動を示したり、アメーバ状の運動をすることは周知のことですが、これは形態、機能ともにアメーバと同じだといえます。生体内のアメーバといえるでしょう。
③ 共生現象
『共生』という語は1879年にバリーが用いたもので、2種類の生物が常に密接な形態的、生理的関係を保ち、また互いに利益を与えあって永続的な共存共栄の生活をしている現象をいいます。
バリーはその最もよい例として地衣類の生活を挙げています。一般に共生現象は案外、割り切った考えで見過ごされているものも少なくないようですが、詳細に研究すると共生と共食生活、そして寄生との間に区別をつけることが困難な場合もあります。場合によっては共生を広義に解して共食生活を含ませ、また相互に生理的な意義をもたないで共に生活している場合を含ませる場合もあります。 生物界を広く、そして深く観察するとき、前述したような共生現象は動的なものであり、時間の経過とともに共生関係が一見すると寄生ではないかと思われる関係もあり、しかもその実は2種の生物の融合によって新しい細胞や新種の生物に進化することもあることは、これまで余り知られていませんでした。ここではこの重要な意義をもつ共生現象と細胞の起源ついて概略を述べておきましょう。
(1) 共食生活
これは『食卓を共にする』という語からきています。2種の生物が互いに食べ物を分けあって食べつつ生活する場合をいいます。この例としてヤドカリとイソギンチャクとの関係を範疇に含めたり、スズメの一種がアフリカのサイやゾウの背にとまってその皮膚に寄生している害虫を喰う例、またこれに似た現象として英国のムクドリとヒツジとの関係も挙げられます。
また海中生物ではある種のカイメン内部には小エビが隠れて生活していたり、サメとコバンザメの関係、さらにある種のカイメン体内には二万種近い他種生物が生活しているのを観察した学者は、このカイメンを『生活用ホテル』…Living hotel…と命名しています。千島も長良川産の淡水カイメン体内に数十種の微生物が生活しているのを観察しています。このような例を共生生活と呼んでいますが、多くの場合が偏利生活(共生者の一方だけが利益を得ている生活)だといえそうです。
(2) 共生
この語源は『共に生きる』の意からきた術語であり、自然界においては共生と共食生活との間に厳密な境界を設けることは不可能でしょう。両者の間には中間的なゾーンがあるのが当然だからです。
寄生虫と宿主とが長い進化史のなかで相互に抵抗性と適応性を獲得し一定の平衡関係を保って共存し、外観的には宿主生物が大きな被害を受けることもなく健康を保持している場合もあります。
こんな場合は寄生と共生との間に明解な限界を引くことはできません。
共生現象は2種生物中の一方が他の生物個体の組織中或いは細胞内に入り込んでいる場合を内部共生、個体の外側に付着するものを外部共生といいこの二つに分けられています。
後者の例としてはアメリカ産のアリが、植物の葉を巣に運び込み堆積させその中に生ずる菌糸を食べて生活する例や、白アリが排泄物で作った団子のなかに生ずる菌糸を食べて生活する例がありますが、このような共生は2種の生物体の細胞や本質的な働きには直接の関係は余りないようです。
一方前者の内部共生は細胞の成長、進化などと直接に重要な関係をもっています。これについては機会があったときお話しましょう。
④ 地衣類における共生現象
すべての地衣類は菌類と藻類との共生によって生じ、地衣類の白色の部分は菌糸でこれが緑顆層を構成しています。菌類の芽胞は発芽しますがその若い葉状体は、適当な藻類との遭遇がなければ発育を中止してしまいます。これは藻類が炭素同化作用によって有機物を合成し菌に与え、菌は水分及び水に溶解している無機塩類を吸収しその一部を藻類に与えるという共生生活ができないからです。
地衣類の種類によって共生する菌と藻の種類は大体が一定ですが時には、同一の菌が種々異なる他種の藻類と適応して共生し、別種の地衣類を形成することもあるという報告があります。
2種の地衣類が同一葉状体のように並んで生育し、或いは反対に同一藻類が種々な菌糸とともに別の地衣類を形成することも知られています。また地衣類は菌糸を従来のものと取り替えることにより他種の地衣類に種の転換を起こすこともあります。
地衣類を構成している菌類と藻類が共生体の特性だとされる『相互に等しい利益を受けあっている』か否かは、一般に考えられている程はっきりしてはいません。今日においても、次の4説があって未解決のまま残されています。
① 菌類は藻類を喰って生きていて一種の寄生性がうかがえる。
② 藻類が菌類に寄生している。
③ 両者は互いに共生生活をしている。
④ 同等の利益交換はない。
①説を主張しているスゥエンドナーは菌類が主人であり、藻類は従者だといっています。クモがその巣で虫を捕らえるように、菌類が藻類を取り囲み、その栄養を吸い取るため藻類は数代後にはもはやその存在が認められなくなるといっています。ウオーニングも藻類は独立の生活を営むことができるが、菌類は藻類を必要とします。だから菌類と藻類は特殊な寄生であるとし、ダニロフも菌糸は藻類の内部に侵入して内容を吸収し藻類を死に至らしめるから寄生だと考えています。
②説は①説とは反対に藻類こそ寄生性をもち菌類は宿主だとするものです。ベジェリックは地衣類から分離した藻類を硝酸と糖分とで培養を試みましたが成功しませんでしたが、有機性のペプトンを加えたところ始めて培養に成功しました。このことから藻類は菌類から栄養を受けているから藻類こそ寄生性のものだと主張しました。
③説は前2説の中間説でバリーやリンクたちによって支持されている説です。リンクは藻類と菌類との関係は緑色植物における葉と根の関係に等しいもの、即ち藻類は自養性で空気中の炭酸ガスから炭水化物を光合成し、菌類からはナトリウムやアルブミン、水分、無機物などを与えられます。
菌類は藻類の合成した炭水化物の助けをかりてアルブミンその他の物質を合成するから両者は正に共生であると主張しています。
④説はコーレイの主張する説で、菌類と藻類とは地衣類において相互に密接に適応していて独立性は失っていますがそのような場合でも藻類は菌類よりも独立生活をしやすい性質をもっています。
コダッツやその弟子は研究結果から『藻類がペプトンを要求するということで、必ずしもそれを寄生虫のものだと判断することはできない。なぜなら他の下等藻類でも同様の性質をもっているから……。有機的栄養分を要求することは場合によって菌類にも藻類にもある。要するに地衣類における菌類と藻類との関係は長い間の相互適応の結果生じたものであり、両種生物はその結果、本来の性質が変更され最初の特性を失ったものである』と述べています。
前述した4つの説に対し、千島喜久男は次のような意見を述べています。
『私は地衣類を深く研究したことはないが、菌類その他についてはこれまでの自己の研究を総合して判断するとき、この4つの説は何れも一面の真理を語るものではあるが、どれも不十分な点があると考える。まず第一に共生現象はこれまでかなり固定的に考えられがちだった。なるほど地衣類の生活段階の一定時期においては、菌と藻は互いに等しい利益を交換している時もあっただろう。いわゆる定型的な共生の時代がそれである。…①説…しかし、環境の変化や時間の経過によって一方が他方を消費することで何れかが比較的優位を示すこともあるだろう。それは①や②説に該当する。
④説はかなり動的な見解であり、私の考えに近いものだが、コーレイは他の一般生物学者と等しく菌類と藻類とはまったく別種のものであるとしている。たしかにそれは今日の生物学的常識からはもっともなことである。しかし、『種』の概念には今日まだ多くの疑義が残されており、地衣類それ自体が種の転換を起こす場合もあるという説も唱えられている。そのうえ、菌類と藻類との関係は菌糸と胞子との関係に似ている。現に私はある種のカビにおいて、菌糸から生じたいわゆる胞子は藻類と近似的なものではないかと推測している。菌糸が膨大して生じた胞子または芽胞様体が地衣類の組織内では、いわゆる藻類になるのではないかということも推測できる。『菌糸が藻類を取り巻き或いはその内部に侵入している状態』と考えられているものは実は、菌糸の一端から藻類を形成しつつある状態だと考えられないこともない。地衣類において菌類と藻類が常に共存している理由を、両者が偶然に遭遇した結果であると考えるより、一定の環境条件と基質の存在とによって、その場所にバクテリアの発生を先駆として菌類や藻類の発生が可能であろうと私は考える。コーレイの主張する菌類と藻類とを純粋培養することがこの問題の解決策だという考えには賛同できない。なぜなら、地衣類の生育する自然的環境とはまったく異なる人工培地での実験結果は、この場合、正しい意味をもたないことになるからである。
第二には、現代生物学では生物の発生、成長、進化をすべて細胞分裂を基調としているが、地衣類の場合、細胞分裂がその発生、成長、増殖の基礎となっいることを実証した人があることを私は知らない。恐らくそれは不可能であろう。私の主張している微生物の新生と、それらの融合と分化による新しい細胞、個体、新種への発展進化という観方からすれば、地衣類を構成する2種の生物の関係は時に共生と見られ、時には寄生と考えられる面があっても、それらは凡て時間、環境の諸条件を通じて生物が下次段階の単位からより高次の生物に進化する動的変化の一つの相を示しているものと解するのが妥当だろう。』
